 Ý Ý
    Ý Ý
   |   Ý
Ý
|
|
|
| Ý |
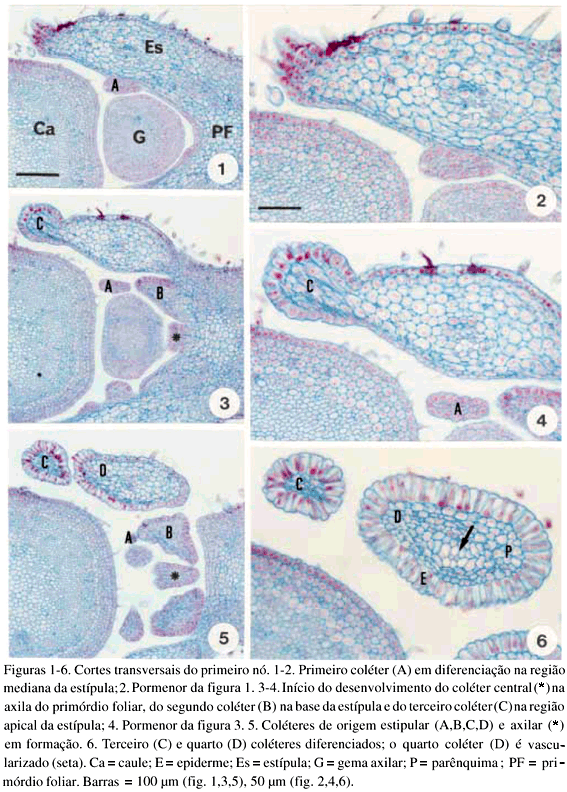
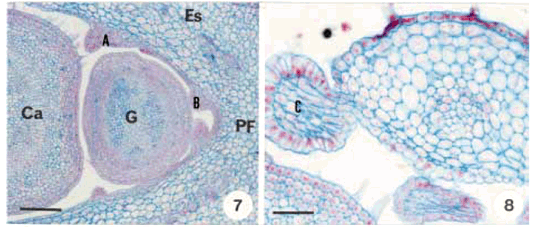
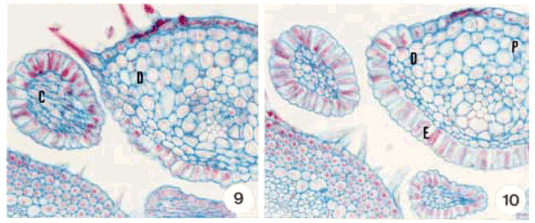
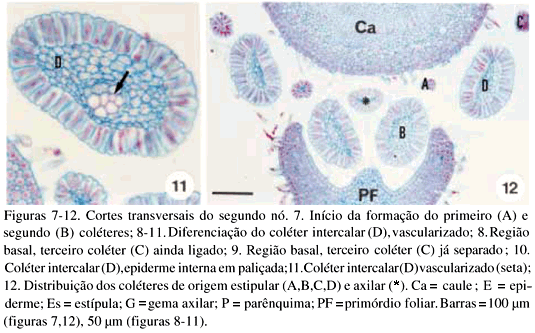
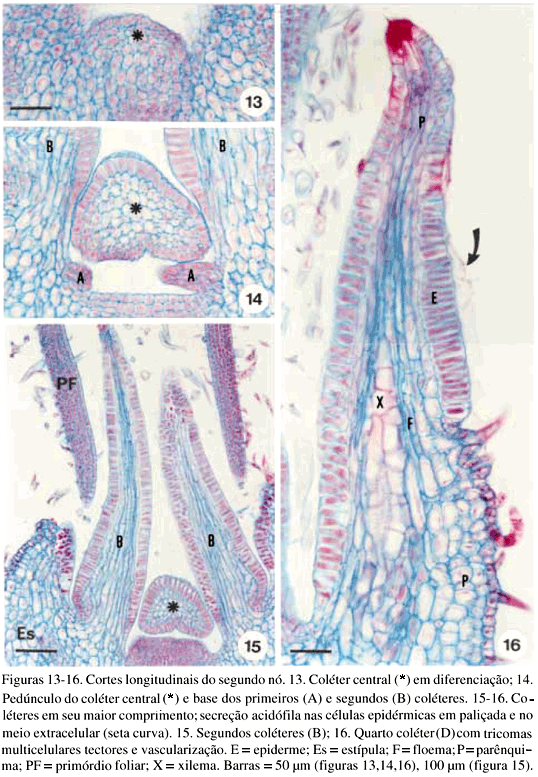
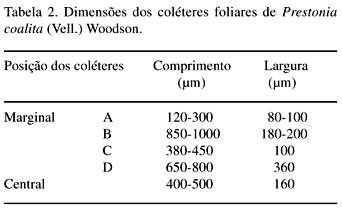
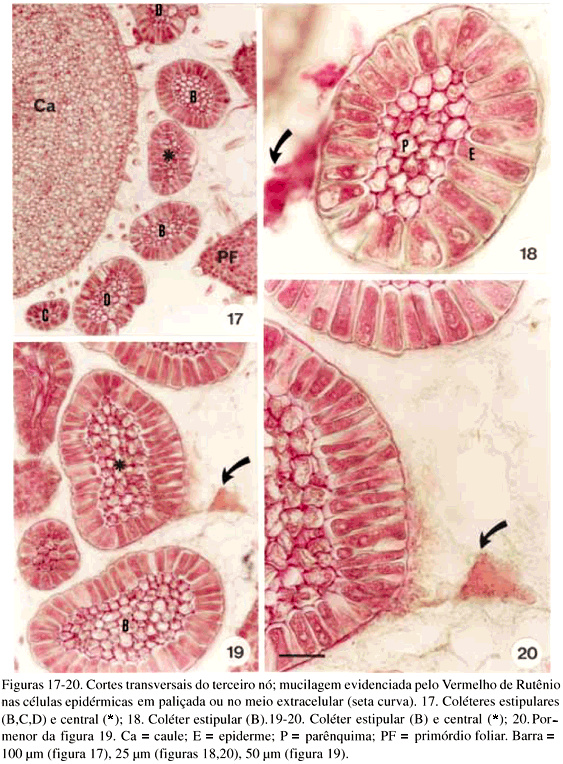
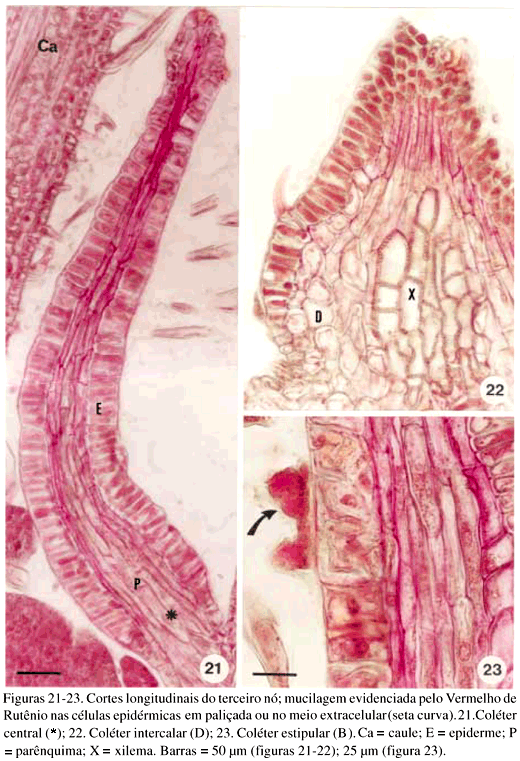
DistribuiÁ„o e caracterizaÁ„o anatÙmica dos colÈteres foliares de Prestonia coalita (Vell.) Woodson (Apocynaceae)1 Ý MARIA C. SCATOLIN RIO2, MARÕLIA M. CASTRO2, 3 e LUIZA S. KINOSHITA2 Ý (recebido: 17 de outubro de 2001; aceito: 29 de maio de 2002) Ý Ý ABSTRACT – (Distribution and anatomical characterization on the foliar colleters of Prestonia coalita (Vell.) Woodson (Apocynaceae)). A structural study on the foliar colleters of Prestonia coalita (Vell.) Woodson was conducted in order to establish their number and positions, to describe their structure, to determine in which stage of leaf development secretion starts and to comprove the presence of mucilage in its secretion. Each node presented 14-18 intrapetiolar colleters, whose differentiation occurs in the early stages of leaf development, and secretion occurs in leaf primordia with 5-8 mm length. Colleters are constituted by a long head that includes a central core of parenchymatous cells, surrounded by one layered palisade secretory epidermal cells, and a short stalk devoid of any epidermal secretory cells. Multicellular hairs and vascular tissue were observed only in intermediary colleters, which correspond with modified stipules.These structures' identification as colleters is confirmed by the presence of mucilage in secretion. The stipular nature of most colleters is confirmed, while only central colleters are originated from leaf axil. RESUMO – (DistribuiÁ„o e caracterizaÁ„o anatÙmica dos colÈteres foliares de Prestonia coalita (Vell.) Woodson (Apocynaceae)). Um estudo estrutural dos colÈteres foliares de Prestonia coalita (Vell.) Woodson foi realizado com a finalidade de definir o n˙mero e a posiÁ„o ocupada pelos colÈteres, caracterizar a sua estrutura, determinar o est·dio de desenvolvimento da folha em que eles se encontrem em fase secretora e comprovar a presenÁa de mucilagem na secreÁ„o por eles produzida. Em cada nÛ, 14-18 colÈteres intrapeciolares s„o observados; sua diferenciaÁ„o ocorre nos est·dios iniciais do desenvolvimento foliar e a atividade secretora em primÛrdios com ca. 5-8 mm de comprimento. Os colÈteres s„o constituÌdos por uma longa cabeÁa, formada por um n˙cleo central de cÈlulas parenquim·ticas rodeado por epiderme uniestratificada secretora em paliÁada, e um curto ped˙nculo, no qual as cÈlulas epidÈrmicas n„o secretoras apresentam formato c˙bico a retangular. Tricomas multicelulares tectores e tecido vascular est„o presentes apenas nos colÈteres intercalares, que correspondem ýs estÌpulas modificadas. A identificac„o destas estruturas como colÈteres È comprovada devido ý presenÁa de mucilagem na secreÁ„o. A natureza estipular da maioria dos colÈteres È confirmada, enquanto apenas os centrais s„o originados na regi„o axilar da folha. Ý Key words - Colleters, leaves, secretion, Prestonia coalita, Apocynaceae Ý Ý IntroduÁ„o ColÈteres s„o estruturas secretoras que produzem uma subst‚ncia viscosa, constituÌda por uma mistura de mucilagem e terpenos, que lubrifica e protege gemas em inÌcio de desenvolvimento (Fahn 1979). Estruturalmente, eles podem ser tricomas, emergÍncias ou gl‚ndulas, neste caso, se vascularizadas (Fahn 1990). Eles podem estar presentes em Ûrg„os vegetativos, sendo mais comuns em gemas e folhas jovens do que em Ûrg„os adultos, e/ou reprodutivos de diversas famÌlias de dicotiledÙneas (Fahn 1979, Thomas 1991). Devido ýs semelhanÁas quanto ý estrutura e distribuiÁ„o dos colÈteres encontrados em partes vegetativas, eles podem freq¸entemente ser confundidos com nect·rios extraflorais (Dave & Patel 1975, Mohan & Inamdar 1986, Thomas 1991). Somente atravÈs da realizaÁ„o de testes quÌmicos, os principais componentes do material secretado s„o determinados e a estrutura que os produz È identificada; desta forma, a presenÁa de aÁ˙car na secreÁ„o dos nect·rios extraflorais os diferencia dos colÈteres, nos quais a mucilagem predomina na secreÁ„o (Fahn 1979). No caso das Apocynaceae s.l., uma famÌlia composta por 424 gÍneros distribuÌdos em cinco subfamÌlias (Endress & Bruyns 2000), os colÈteres presentes nas partes vegetativas constituem-se em emergÍncias ou gl‚ndulas persistentes observadas na base da l‚mina foliar e na face adaxial da base do pecÌolo (Thomas 1991), podendo ocupar posiÁ„o interpeciolar ou intrapeciolar (Sennblad et al. 1998). Embora os colÈteres sejam estruturas freq¸entes nas Apocynaceae, com ocorrÍncia em folhas registrada para 50 gÍneros (tabela 1), poucos trabalhos relacionados ý estrutura, distribuiÁ„o, desenvolvimento e/ou histoquÌmica dos colÈteres de espÈcies dessa famÌlia foram realizados nos ˙ltimos anos, destacandose aqueles efetuados em Aganosma caryophyllata G. Don. (Dave et al. 1987), Allamanda cathartica L., Alstonia scholaris L., Nerium indicum Mill. (Thomas & Dave 1989a, b, c), Roupelia grata Wall. (hoje Strophanthus gratus (Wall. et Hook.) Baill. et Arn.; Thomas et al. 1989), Mandevilla illustris (Vell.) Woodson e M. velutina (Mart. ex Stadelm.) Woodson (Appezzato-da-GlÛria & Estelita 2000). Podem, ainda, ser mencionados os trabalhos relacionados ý ontogÍnese dos colÈteres em Allamanda cathartica e Tabernaemontana divaricata (L.) R. Br. (Ramayya & Bahadur 1968), ao modo de secreÁ„o de colÈteres em Alstonia scholaris (Thomas & Dave 1990) e aos aspectos filogenÈticos de colÈteres em Apocynaceae (Thomas & Dave 1991). Ý Ý O gÍnero Prestonia R. Br. pertence ý subfamÌlia Apocynoideae Burnett (Endress & Bruyns 2000) e È constituÌdo por cerca de 60 espÈcies de lianas com distribuiÁ„o neotropical, onde podem ser encontrados colÈteres na base dos pecÌolos e na base dos lobos do c·lice (Woodson 1936). Segundo Rio (2001), os colÈteres interpeciolares presentes nos nÛs vegetativos de algumas espÈcies de Prestonia tÍm import‚ncia taxonÙmica, sendo utilizados em chave para identificaÁ„o das espÈcies ocorrentes nas regiıes Sul e Sudeste do Brasil. Uma vez que n„o h· estudos anatÙmicos referentes aos colÈteres encontrados nas espÈcies de Prestonia, a presente investigaÁ„o teve por objetivo definir o n˙mero e a posiÁ„o ocupada pelos colÈteres foliares de Prestonia coalita (Vell.) Woodson, caracterizar a sua estrutura, determinar o est·dio de desenvolvimento da folha em que os colÈteres se encontrem em fase secretora e comprovar a presenÁa de mucilagem na secreÁ„o produzida por estas estruturas. Ý Material e mÈtodos Para o estudo dos colÈteres foliares, um indivÌduo de Prestonia coalita (Vell.) Woodson foi coletado em Carrancas, MG.O material testemunha est· depositado no Herb·rio da Universidade Estadual de Campinas (UEC): BRASIL: MINAS GERAIS: Carrancas, 24/II/2000, L.S. Kinoshita et al. 2000.10 (UEC). O material foi fixado em FAA 50 por 48 h. e estocado em ·lcool etÌlico 70% (Johansen 1940). Os ·pices e dois nÛs subseq¸entes foram isolados a partir dos ramos vegetativos. As peÁas foram desidratadas pela sÈrie butÌlica (Johansen 1940) e incluÌdas em paraplast. Os cortes seriados longitudinais e transversais, com 12 mm de espessura, foram corados para o estudo estrutural com Safranina (soluÁ„o alcoÛlica) 1% e Azul de Astra (soluÁ„o aquosa) 1% (Gerlach 1969) e as l‚minas permanentes montadas em resina sintÈtica. Para evidenciar a presenÁa de mucilagem (Gregory & Baas 1989), os cortes foram corados com Vermelho de RutÍnio (soluÁ„o aquosa) 1:5.000 (Rawlins & Takahashi 1952) e as l‚minas semipermanentes, montadas em gelatina glicerinada. As fotomicrografias foram obtidas em microscÛpio Olympus BX 50, com a utilizaÁ„o de filme Kodak Gold 35Ýmm asa 100. As escalas foram obtidas atravÈs da projeÁ„o de l‚mina micromÈtrica nas mesmas condiÁıes Ûpticas utilizadas para as ilustraÁıes. Ý Resultados Na regi„o nodal do caule de Prestonia coalita, um n˙mero vari·vel de colÈteres È observado na base da folha ocupando posiÁ„o intrapeciolar; estes colÈteres tÍm origens independentes e s„o formados no inÌcio do desenvolvimento dos primÛrdios foliares (figuras 1-15), que apresentam filotaxia oposta. Em cada nÛ, 14-18 colÈteres podem ser encontrados, sete a nove por primÛrdio, dos quais apenas um ocupa posiÁ„o axilar e os demais posiÁ„o marginal (figuras 5, 12). Ý Ý Ý Ý Ý Ý O mesmo padr„o de desenvolvimento dos colÈteres se repete em todos os nÛs estudados; assim, a descriÁ„o destas estruturas ser· efetuada considerando-se o primeiro e segundo nÛ vegetativo (figuras 1-16) de apenas um lado correspondente ý base de um dos dois primÛrdios foliares opostos, onde podem ser encontrados nove colÈteres por nÛ. A origem de oito (quatro de cada lado) deles se d· na regi„o marginal do primÛrdio foliar correspondente ý estÌpula (figuras 15), e de um (o central), na regi„o axilar do primÛrdio foliar (asterisco; figuras 3, 13-14). Os dois primeiros colÈteres (A) comeÁam a se desenvolver na regi„o mediana da estÌpula (figuras 1-4, 7-8, 14), estando ausentes nas folhas com apenas sete colÈteres. Os segundos (B) tÍm origem na regi„o basal da estÌpula (figuras 3, 7, 14-15), enquanto os terceiros (C) s„o formados na regi„o apical (figuras 3-6, 8). Os quartos colÈteres (D) ocupam posiÁ„o intercalar entre os segundos (B) e terceiros (C) colÈteres e mantÍm a vascularizaÁ„o das estÌpulas (figuras 3-6, 8-12, 16), diferenciando-se, portanto, dos demais colÈteres que n„o s„o vascularizados (figura 15). Observa-se a manutenÁ„o do feixe vascular durante a diferenciaÁ„o desses colÈteres (figuras 3-6, 8-11). As dimensıes dos colÈteres foliares de P. coalita est„o descriminadas na tabela 2. Ý Ý Todos os colÈteres foliares s„o constituÌdos por uma cabeÁa longa, formada por um n˙cleo central de cÈlulas parenquim·ticas rodeado por uma epiderme secretora uniestratificada em paliÁada, sobre um ped˙nculo curto, no qual as cÈlulas epidÈrmicas n„o secretoras tÍm formato c˙bico a retangular. A cabeÁa apresenta um formato cÙnico, pontiagudo no ·pice, enquanto o ped˙nculo pode estar reduzido n„o apresentando estreitamento (figuras 15-16, 21). Todo o colÈter È recoberto por uma fina cutÌcula e o tecido vascular est· presente apenas nos quartos colÈteres (D), que ocupam posiÁ„o intercalar; nestes colÈteres, tricomas multicelulares tectores tambÈm foram observados (figuras 16, 22), sendo os ˙nicos dentre todos os colÈteres foliares que apresentaram modificaÁıes estruturais. As cÈlulas epidÈrmicas em paliÁada da cabeÁa do colÈter, respons·veis pela secreÁ„o do exsudato, tÍm paredes celulares finas, n˙cleo relativamente grande e citoplasma denso, caracterÌsticas comuns a cÈlulas secretoras (figuras 10-11, 16). Uma secreÁ„o acidÛfila foi evidenciada pela Safranina tanto no interior das cÈlulas epidÈrmicas quanto no meio extracelular (seta curva; figura 16) de colÈteres do segundo nÛ, nos quais cÈlulas com citoplasma de aspecto denso e outras com retraÁ„o do protoplasto tambÈm podem ser observadas (figuras 10-11, 16). Ý Ý Ý Ý A presenÁa de mucilagem foi evidenciada no meio extracelular (seta curva; figuras 18-20, 23) e no interior das cÈlulas epidÈrmicas em paliÁada que recobrem a cabeÁa de todos os colÈteres observados (figuras 17-23), tanto no segundo (primÛrdios foliares com ca. 5 mm de comprimento) quanto no terceiro nÛ vegetativo (primÛrdios foliares com ca. 8 mm). Essas cÈlulas secretoras apresentam citoplasma com conte˙do de aspecto denso ou, ent„o, retraÁ„o do protoplasto (figuras 18-20), sinal de liberaÁ„o da secreÁ„o; porÈm, o rompimento da cutÌcula n„o foi observado. Em corte longitudinal (figura 21), o material produzido pelas cÈlulas secretoras, que ocupam as regiıes apical e mediana da cabeÁa dos colÈteres, ainda se encontra em seu interior; tais cÈlulas n„o apresentam retraÁ„o do citoplasma, como as cÈlulas da regi„o basal da cabeÁa do colÈter. N„o h· indÌcios de atividade secretora nas cÈlulas do ped˙nculo (figura 21). Ý Discuss„o ColÈteres em folhas j· foram observados em espÈcies pertencentes a 19 gÍneros da subfamÌlia Apocynoideae, com duas ocorrÍncias para l‚mina foliar em Forsteronia e Mandevilla, e as demais para pecÌolo em espÈcies de Adenium, Aganosma, Apocynum, Baissea, Beaumontia, Forsteronia, Holarrhena, Ichnocarpus, Mandevilla, Nerium, Parsonsia, Prestonia, Stephanostema, Strophanthus, Thenardia, Trachelospermum, Urceola, Vallaris e Wrightia (Hansen 1985, Dave et al. 1987, Thomas et al. 1989, Thomas & Dave 1989c, 1991, Thomas 1991, Sennblad et al. 1998, Appezzato-da-GlÛria & Estelita 2000). Em Prestonia, os colÈteres foliares foram observados e macromorfologicamente descritos para as 15 espÈcies ocorrentes nas regiıes Sul e Sudeste do Brasil (RioÝ2001). Em Prestonia coalita, cada folha possui de sete a nove colÈteres intrapeciolares. VariaÁıes no n˙mero de colÈteres nas folhas de uma mesma espÈcie j· foram registradas para a famÌlia. Ramayya & Bahadur (1968) observaram 10-13 colÈteres foliares em Allamanda cathartica, enquanto Thomas & Dave (1989a) registraram apenas quatro a seis colÈteres foliares para esta espÈcie. Em Thevetia peruviana (Pers.) K. Schum (= T. neriifolia Juss.), Hifny Saber et al. (1969 apud Thomas 1991) mencionaram oito a dez colÈteres peciolares e Fjell (1983) registrou 14-18 colÈteres peciolares para esta espÈcie. Segundo Thomas (1991), o n˙mero dos colÈteres de uma espÈcie pode variar com a distribuiÁ„o geogr·fica. Embora o n˙mero e a posiÁ„o dos colÈteres j· tenham sido referidos como caracteres taxonÙmicos relevantes para as Apocynaceae (Woodson & Moore 1938), tais aspectos tÍm sido pouco considerados nos estudos efetuados em partes vegetativas. A presenÁa de colÈteres interpeciolares foi uma caracterÌstica utilizada na chave para a identificaÁ„o das 15 espÈcies de Prestonia ocorrentes nas regiıes Sul e Sudeste do Brasil (Rio 2001). P. solanifolia (M¸ll. Arg.) Woodson foi diferenciada de P. coalita e de P. dusenii (Malme) Woodson, espÈcies intimamente relacionadas quanto ý morfologia floral, assim como P. calycina M¸ll. Arg.foi separada das demais espÈcies com indumento abundante nas folhas, por apresentarem colÈteres interpeciolares. Da mesma forma, P. perplexa Woodson e P. denticulata (Vell.) Woodson foram distintas das demais espÈcies com apÍndices supra-estaminais e folhas glabras a levemente papiladas pela presenÁa de colÈteres interpeciolares, sendo posteriormente diferenciadas por caracteres foliares e florais. Os colÈteres foliares de Prestonia coalita s„o do tipo padr„o ("standard" – S) descrito por Lestern (1974), caracterizado pela presenÁa de uma cabeÁa alongada recoberta por epiderme secretora em paliÁada e um ped˙nculo curto sem cÈlulas secretoras. Apesar de ter sido proposta para colÈteres encontrados em espÈcies de Rubiaceae, a terminologia definida por Lestern (1974) tem sido utilizada para a descriÁ„o dos colÈteres de diversas famÌlias, inclusive Apocynaceae (Thomas 1991, Appezzato-da-GlÛria & Estelita 2000). Quanto ý presenÁa de tecido vascular, somente os colÈteres intercalares de P. coalita s„o vascularizados por reterem a vascularizaÁ„o da estÌpula; os demais colÈteres, inclusive os de maior tamanho, n„o s„o vascularizados. Este È o primeiro registro de colÈteres vascularizados para Prestonia. ColÈteres foliares vascularizados ocorrem em Aganosma caryophyllata (Dave et al. 1987), Mandevilla illustris e M. velutina (Appezzato-da-GlÛria & Estelita 2000). Os colÈteres de Nerium oleander L. (Williams et al. 1982), Allamanda cathartica (Thomas & Dave 1989a) e Roupelia grata (Thomas et al. 1989) n„o apresentaram tecido vascular. Ressalta-se a import‚ncia de analisar os colÈteres atravÈs de cortes seriados transversais e longitudinais, tanto para a constataÁ„o da ocorrÍncia de vascularizaÁ„o, que dependendo do plano do corte pode ou n„o estar evidente (Appezzato-da-GlÛria & Estelita 2000), quanto para a definiÁ„o do n˙mero real de colÈteres encontrado por peÁa. Dentre as modificaÁıes estruturais (presenÁa de tricomas, cÈlulas subepidÈrmicas procumbentes e laticÌferos) j· registradas para colÈteres da famÌlia Apocynaceae (Thomas 1991), apenas tricomas multicelulares tectores foram encontrados exclusivamente nos colÈteres intercalares derivados diretamente da estÌpula de Prestonia coalita. Tricomas unicelulares j· foram registrados para Aganosma caryophyllata (Dave et al. 1987), enquanto Roupelia grata (Thomas et al. 1989) e Nerium indicum (Thomas & Dave 1989c) apresentaram uma camada de cÈlulas subepidÈrmicas alongadas radialmente. Apenas laticÌferos foram encontrados em Allamanda cathartica (Thomas & Dave 1989a). Thomas & Dave (1991) consideraram as modificaÁıes estruturais, assim como a vascularizaÁ„o, representantes de um passo evolutivo entre os colÈteres de Apocynaceae. Em cada folha de Prestonia coalita, um ˙nico colÈter tem origem na regi„o axilar do primÛrdio foliar, enquanto que os demais s„o originados a partir de expansıes laterais da base do primÛrdio foliar (estÌpulas). Dessa forma, a natureza estipular È confirmada para a maioria dos colÈteres foliares de P. coalita. Woodson & Moore (1938) propuseram a teoria estipular para os colÈteres de Apocynaceae, considerando que tanto os colÈteres dos nÛs vegetativos quanto os calicinais pertenceriam ý categoria de estÌpulas. Considerando a presenÁa de colÈteres axilares em P. coalita, o estudo dos colÈteres nodais deveria ser estendido para outras espÈcies de Apocynaceae com o propÛsito de reavaliar a teoria proposta por Woodson & Moore (1938). A retenÁ„o dos colÈteres com tendÍncia ý reduÁ„o das estÌpulas foi observada em todas as famÌlias da ordem Gentianales (Thomas 1991). A secreÁ„o dos colÈteres de Prestonia coalita pode ser encontrada em est·dios muito iniciais do desenvolvimento da folha (primÛrdios de 5 a 8 mm), sendo constituÌda por mucilagem. Segundo Thomas (1991), os colÈteres comeÁam a secretar antes da expans„o da l‚mina foliar, o que foi confirmado em P.Ýcoalita. A natureza mucilaginosa da secreÁ„o e a fase inicial do desenvolvimento foliar em que os colÈteres s„o encontrados em atividade secretora est„o relacionadas ý funÁ„o que essas estruturas desempenham, que È a de lubrificar e proteger gemas e, conseq¸entemente, meristemas em inÌcio de desenvolvimento (Fahn 1979). Ý Agradecimentos – Os autores agradecem ý FundaÁ„o de Amparo ý Pesquisa do Estado de S„o Paulo (Fapesp) pelas bolsas concedidas (proc. n. 96/10106-3, 99/01768-0) e ao Fundo de Apoio ý Pesquisa da Universidade Estadual de Campinas (FAEP/Unicamp) pelo auxÌlio ý publicaÁ„o das pranchas coloridas (sol. n. 0612/02). Ý ReferÍncias bibliogr·ficas APPEZZATO-DA-GL”RIA, B. & ESTELITA, M.E.M. 2000. Development, structure and distribution of colleters in Mandevilla illustris e M. velutina (Apocynaceae). Revista Brasileira de Bot‚nica 23:113-120.
DAVE, Y.S. & PATEL, N.D. 1975. A developmental study of extrafloral nectaries in slipper spurge (Pedilanthus tithymaloides, Euphorbiaceae). American Journal of Botany 62:808-812. DAVE, Y., THOMAS, V. & KURIACHEN, P.M. 1987. Structure and development of colleters of Aganosma caryophyllata G. Don. Pakistan Journal of Botany 19:243-248. ENDRESS, M.E. & BRUYNS, P.V. 2000. A revised classification of Apocynaceae s.l. The Botanical Review 66:1-56. FAHN, A. 1979. Secretory tissues in plants. Academic Press Inc., London. FAHN, A. 1990. Plant anatomy. 4th ed. Pergamon Press, Oxford. FJELL, I. 1983. Anatomy of the xeromorphic leaves of Allamanda neriifolia, Thevetia peruviana and Vinca minor (Apocynaceae). Nordic Journal of Botany 3:383-392. GERLACH, G. 1969. Botanische mikrotechnik, eine linfuhrung. Georg Thieme, Stuttgard. GREGORY, M. & BAAS, P. 1989. A survey of mucilage cells in vegetative organs of the dicotyledons. Israel Journal of Botany 38:125-174. HANSEN, B.F. 1985. A monographic revision of Forsteronia (Apocynaceae). PhD Thesis, University of South Florida, Florida. JOHANSEN, D.A. 1940. Plant microtechnique. McGraw-Hill Book Co., New York. LESTERN, N.R. 1974. Morphology and distribution of colleters and cristals in relation to the taxonomy and bacterial leaf nodule symbiosis of Psychotria (Rubiaceae). American Journal of Botany 61:973-981. MOHAN, J.S.S. & INAMDAR, J.A. 1986. Ultrastructure and secretion of the extrafloral nectaries of Plumeria rubra. Annals of Botany 57:389-401. RAMAYYA, N. & BAHADUR, B. 1968. Morphology of the "squamellae" in the light of their ontogeny. Current Science 18:520-522. RAWLINS, T.E. & TAKAHASHI, W.N. 1952. Technics of plant histochemistry and virology. The National Press, Milbrae. RIO, M.C.S. do 2001. Estudos taxonÙmicos e anatÙmicos do gÍnero Prestonia R. Br. nom. cons. (Apocynaceae). Tese de mestrado, Universidade Estadual de Campinas, Campinas. SENNBLAD, B., ENDRESS, M.E. & BREMER, B. 1998. Morphology and molecular data in phylogenetic fraternity: the tribe Wrightieae (Apocynaceae) revisited. American Journal of Botany 85:1143-1158. THOMAS, V. 1991. Structural, fuctional and phylogenetic aspects of the colleter. Annals of Botany 68:287-305. THOMAS, V. & DAVE, Y. 1989a. Histochemistry and senescence of colleters of Allamanda cathartica L. (Apocynaceae). Annals of Botany64:201-203. THOMAS, V. & DAVE, Y. 1989b. The colleters of Alstonia scholaris L. (Apocynaceae). Indian Botanical Contactor 6:25-29. THOMAS, V. & DAVE, Y. 1989c. Structure, origin, development and senescence of colleters in Nerium indicum Mill. (N. odorum Soland., Apocynaceae). Korean Journal of Botany 32:163-172. THOMAS, V. & DAVE, Y. 1990. Mode of secretion in the colleters of Alstonia scholaris (Apocynaceae). Phyton 30:209-212. THOMAS, V. & DAVE, Y. 1991. Comparative and phylogenetic significance of colleters in Apocynaceae. Feddes Repertorium 102:23-28. THOMAS, V., DAVE, Y. & MENON, A.R.S. 1989. Anatomy and histochemistry of colleters in Roupelia grata (Apocynaceae). Nordic Journal of Botany 8:493-496. WILLIAMS, R.F., METCALFE, R.A. & GUST, L.W. 1982. The genesis of form in oleander (Nerium oleander L. ). Australian Journal of Botany 30:677-687. WOODSON, R.E. 1936. Studies in the Apocynaceae. IV. The American genera of Echitoideae. Annals of the Missouri Botanical Garden 23:169-548. WOODSON, R.E. & MOORE, J.A. 1938. The vascular anatomy and comparative morphology of apocynaceous flowers. Bulletin of the Torrey Botanical Club 65:135-165. Ý Ý 1. Parte da tese de mestrado de Maria Carolina Scatolin do Rio, vinculada ao Programa de PÛs-graduaÁ„o em Biologia Vegetal da Universidade Estadual de Campinas. 2. Universidade Estadual de Campinas, Departamento de Bot‚nica, Instituto de Biologia, Caixa Postal 6109, 13083-970 Campinas, SP, Brasil. 3. Autor para correspondÍncia: mariliac@unicamp.br | Ý |
©Ý2005Ý
SBSP
Caixa Postal 57088
04093-970 S„o Paulo SP - Brasil
Tel.: (55 11) 5584-6300 - ext. 225
Fax: (55 11) 577.3678
rbbot@ig.com.br